American Rose Society, Uncategorized
What Are Stomata Cells?
 by Dr. Gary A. Ritchie • 8026 61st Ave. NE • Olympia, WA 98516
by Dr. Gary A. Ritchie • 8026 61st Ave. NE • Olympia, WA 98516Email: rosedoctor@comcast.netImagine for a moment that you are a nanoperson who can crawl inside a rose leaf. What would it look like? What would it feel like? What sorts of structures would you see?
Above you there would be masses of green, tightly packed cells arranged in palisades. At mid‑leaf, where you are, the cells become loosely packed with large air spaces between them. The surfaces of these cells are wet to the touch. The air is very damp – 100 percent relative humidity in fact. Throughout this labyrinth of cells you see complex systems of pipes running to and from each cell. Some (called xylem) are filled with water, while others (called phloem) are filled with a sugary solution. These are the leaf veins that ultimately connect the cells to the rest of the plant.
Below you is a dark green carpet of thin, tightly interconnected cells – the epidermis – which is covered on its outside by a thin layer of wax called the cuticle. Scattered across the epidermis are thousands of tiny windows leading to the outside world. These windows are the stomata. In previous articles we’ve mentioned stomata in several contexts, but have never discussed them in any detail. Here we will explore what stomata do and how they work.
The main thing stomata do is to open and close. When they are open, gases inside the leaf, such as water vapor, carbon dioxide (CO2) and oxygen, diffuse out of the leaf into the atmosphere. The diffusion of water vapor from the leaf is called transpiration. CO2 that diffuses out is a byproduct of respiration, while the oxygen that escapes is a byproduct of photosynthesis.
Correspondingly, CO2 from outside the leaf can diffuse into the leaf, providing your rose with the carbon it needs to make food. This exchange of gasses is absolutely critical – without stomata plants could not live, life on Earth would cease, and the stock market would tank.
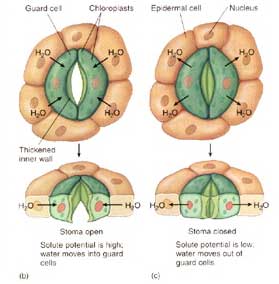
The stomatal pore is not actually a structure in itself. Rather, it is a space created when two cells, called guard cells which are embedded in the epidermis of the leaf, become either turgid (filled with water) or flaccid. The drawing at right shows how this happens. The guard cells are shaped like tiny kidney beans. When they are flaccid there is no space between them – i.e. the stomate (singular of stomata) is closed. But when they are pumped full of water, they stretch and pull apart in the middle, forming a pore.
Ideally, it would be advantageous to plants if stomata were always open. This would allow transpiration and photosynthesis to proceed continuously.
Transpiration is important because it pulls nutrient ions up into the plant, and it cools the leaves on hot days. Photosynthesis is important because it produces food. So, for the plant to gain both nutrients and food the stomata must be open. But this is problematic, because when stomata are open transpiration sucks water from leaves, causing them to dehydrate. When leaves dehydrate, stomata close blocking the uptake of CO2. So the plant must strike a balance between absorbing CO2 while not depleting its internal water balance.
To realize these conflicting objectives, hundreds of millions of years of plant evolution have created an exquisite array of interacting mechanisms and feedbacks. These control and optimize stomatal movement. For example, stomata close in darkness and open in light. The logic for this is clear – photosynthesis requires light. Stomata close when the leaves begin to dry. Again, this is a mechanism that preserves leaf moisture and prevents dehydration. They also close when humidity is low and when the CO2 concentration of the internal leaf atmosphere is high.
Stomatal opening and closing are modulated by what is know as a “potassium pump.” Potassium ions (K+) contained in the guard cells influence their osmotic properties. As the K+ concentration increases, the cell osmotic potential drops. This pulls water into the guard cells, opening the stomata. This active process accumulates potassium ions against a concentration gradient. Therefore, it requires that the plant expend energy.
An amazing discovery was made recently relating stomata to global climate change. Plant physiologists found that stomatal frequency (number of stomata per unit of leaf area) is directly related to the ambient atmospheric CO2 content – as CO2 increases, stomatal frequency decreases. Paleobotanists (kooks who make their living studying plant fossils) can use this relationship to reconstruct ancient CO2 profiles. First, they determine the age of fossil leaves, then they count their stomatal frequency. The more stomata that occur on the leaves, the lower the atmospheric CO2 was when the leaves were formed. This, then, provides a very long-term record of atmospheric CO2 concentrations. Pretty cool stuff.
Other interesting research conducted several decades ago involves indirect stomatal responses to the environment. Scientists found that when soil moisture begins to decline, ever so slightly, around a plant’s roots, the young root tips are able to perceive this stimulus. They then send chemical signals (hormones) up to the leaves that tell the stomata to close in “anticipation” of pending drought.
While this is a very interesting phenomenon, it’s also a great lead‑in to the next series of articles. There we will explore plant hormones – the chemical messengers that move around in plants and tell them what to do and when to do it. You won’t want to miss this, so stay tuned.
This article was provided to the TVRS as a courtesy by the American Rose Society.[/vc_column_text][/vc_column][/vc_row]